





To the Editor:
In cystic fibrosis (CF), repeated inflammation of the lung leads to progressive damage of lung tissue [1, 2]. In case of tissue destruction or in response to inflammation, necrotic cells release danger signals. These proteins trigger inflammatory responses, contribute to elimination of pathogens and induce tissue repair [3]. Imbalanced or uncontrolled release of such mediators may amplify inflammatory responses that in turn facilitate tissue damage.
Interleukin (IL)-33 and high-mobility group box 1 protein (HMGB1) are both considered to be such “alarmins” [3]. Given the T-helper cell (Th)17/Th2 nature of lung inflammation in asymptomatic CF individuals and its linkage to CF-associated pathogens [1, 2], we hypothesised that IL-33 and HMGB1 may contribute to the local sustained inflammatory processes.
Both these alarmins are dual-function proteins, as they are nuclear factors regulating gene expression and also act as cytokines when released from cells [3]. In contrast to ubiquitously expressed HMGB1, IL-33 is mainly expressed in barrier-function-like epithelial cells [3]. IL-33 is mainly linked to augmentation and initiation of Th2-type inflammation but might also antagonise overwhelming tissue inflammation linked to pathogens [4–6]. HMGB1 acts as a cytokine, which contributes to activation of innate receptors, matures dendritic cells (DCs), contributes to migration of DCs and amplifies inflammation itself via induction of pro-inflammatory cytokines like tumour necrosis factor (TNF)-α, IL-6, IL-1β and macrophage inflammatory protein (MIP)1 [7].
To directly compare the contributions of released IL-33 and HMGB1 in CF lung inflammation, we assessed protein levels in bronchoalveolar lavage fluid (BALF) of 44 CF patients and 10 non-CF controls with recurrent pulmonary infections. To increase the homogeneity of the CF population, patients with clinical signs of exacerbation (decline of lung function >10%, start of an antibiotic therapy, increase in coughing, clinical signs of infection, or signs of infection in terms of C-reactive protein elevation or differential counts) were excluded from analysis. In brief, BALF was obtained from patients who underwent bronchoscopy at the CF centre of the Dept of Pediatrics and Adolescent Medicine, Medical University of Vienna (Vienna, Austria) from July 2009 to December 2011, as described by Tiringer et al. [1]. Sampling and processing of BALF and multiplex sandwich ELISA (IL-1β, IL-5, IL-6, IL-8, IL-13, IL-17A, IL-23, interferon (IFN)-γ, thymus and activation-regulated chemokine (TARC), TNF-α, macrophage-derived chemokine (MDC), thymic stromal lymphopoietin (TSLP), IL-17E and IL-12p70) were performed as previously described and correlated with IL-33 and HMGB1 values [1]. IL-33 (eBioscience, San Diego, CA, USA) and HMGB1 (IBL International, Hamburg, Germany) were measured via sandwich ELISA according to the manufacturer’s instructions and absolute values were corrected according to the concentration factor. Diagnosis of CF was confirmed by repeated sweat tests and the type of mutation was defined by genetic testing. Parenchymal and bronchial epithelial tissue samples from end-stage disease patients was obtained at the time of transplantation from explanted lungs at the Dept of Thoracic Surgery, Medical University of Vienna from August 2009 to August 2011. Clinically stable CF patients and controls were matched according to age, sex and body mass index. PCR primers and probes were designed based on the sequences reported in GenBank with the Primer Express software version 1.2 (Applied Biosystems, Foster City, CA, USA) and were as follows: IL-33 forward 5'-GTT ACT TTA GGA GAG AAA CCA CCA AA-3', IL-33 reverse 5'-TTG ACA GGC AGC GAG TAC CA-3', elongation factor (EF)1α forward 5'-CTG AAC CAT CCA GGC CAA AT-3', EF1α reverse 5'-GCC GTG TGG CAA TCC AAT-3'. Relative changes in tissue expression of genes between patient groups were calculated as follows: the sample with the lowest amount of the respective gene was set as internal standard and relative expression was calculated by 2(ΔCt sample of interest-ΔCt lowest sample). Data are expressed as fold expression of the respective sample. The amplification efficacy was initially tested by serial dilution of the cDNA. The generation of relevant primer-dimer formations was excluded.
We found divergent prominence of IL-33 and HMGB1 in local inflammatory processes in CF lungs. Levels of IL-33 were significantly higher in clinically stable CF patients compared with non-CF controls (p<0.05), whereas HMGB1 levels were significantly higher in non-CF controls with recurrent infections (p=0.02) (fig. 1a and b). In CF patients, levels of IL-33 correlated with levels of IL-8 (r=0.51; p<0.01) and IL-13 (r=0.53; p<0.01) and, to a lesser degree, with IL-12p70 (r=0.43; p<0.05), confirming the reported Th2-enhancing and monocyte-recruiting properties of IL-33 and supporting a role in epithelial inflammation via IL-8 [1, 6]. In clinically stable CF patients who were able to perform lung function tests (n=13), levels of IL-33 in BALF negatively correlated with forced vital capacity (r=0.58; p<0.05), suggesting another role for IL-33 apart from pathogen-related infection and tissue destruction. HMGB1 negatively correlated with levels of IL-1β (r= -0.46; p<0.01) [8].
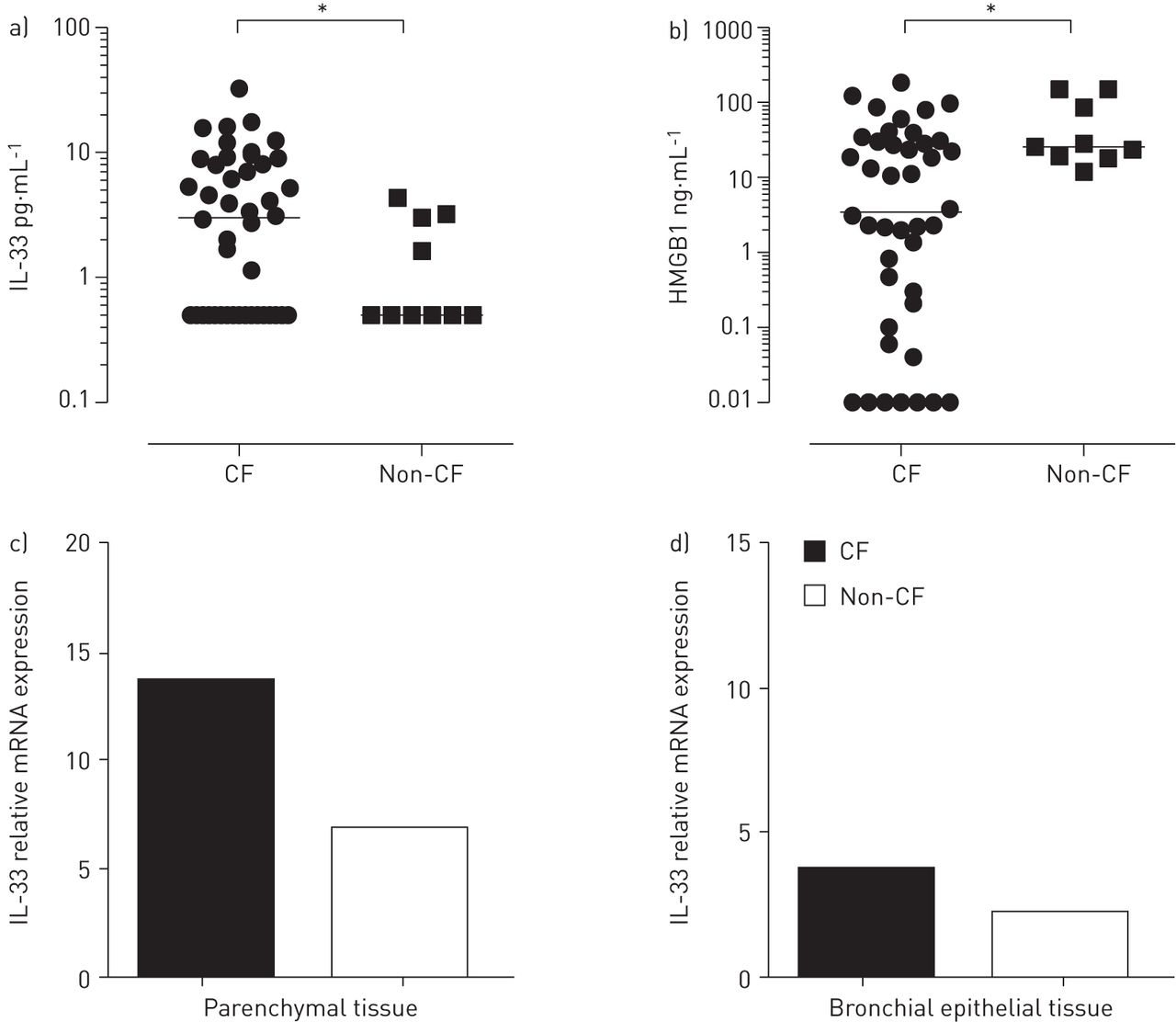
{kind=link}
Interleukin (IL)-33 and high-mobility group box 1 protein (HMGB1) in cystic fibrosis (CF) lungs. Levels of a) IL-33 and b) HMGB1 were measured in bronchoalveolar lavage fluid via ELISA. a) IL-33 was significantly elevated in clinically stable CF patients (n=32) compared to non-CF controls (n=10), whereas b) HMGB1 was higher in the control group (CF group n=32, controls n=9). mRNA expression levels of IL-33 were measured in c) parenchymal and d) bronchial epithelial lung tissue of CF patients (n=9) and non-CF controls (primary pulmonary hypertension and lung fibrosis, n=11) via real-time RT-PCR. The sample with the lowest amount of the respective gene was set as internal standard and relative expression was calculated by 2(ΔCt sample of interest-ΔCt lowest sample). Data are expressed as fold expression of the respective sample. *: p<0.05, Mann–Whitney U-test.
In addition, we assessed expression levels of IL-33 in parenchymal and bronchial epithelial lung tissue from explanted end-stage lungs of nine CF patients and 11 controls (lung fibrosis and primary pulmonary hypertension) via real-time RT-PCR [1]. In line with the aforementioned results, IL-33 mRNA expression was higher in CF patients compared with non-CF controls (fig. 1c and d), although this did not reach statistical significance.
A role for HMGB1 in CF background inflammation and leukocyte recruitment has been described [9]. It may appear surprising that HMGB1 is lower in BALF of CF patients compared with non-CF patients with recurrent infections. However, compared with reported values in healthy controls (1–8 ng·mL−1), it is elevated [2]. Moreover, the control group used by Rowe et al. [9] consisted of healthy individuals, compared with the clinically stable individuals with recurrent pulmonary infections in this study. In addition, binding of HMGB1 to bacterial compounds is reported and greater abundance of such components in the lungs of CF patients compared with non-CF controls seems feasible [7]. The latter may explain the inverse relationship between IL-1β and HMGB1 levels in BALF. Furthermore, cholinergic agonists, which are part of the CF basic inhalant therapy, inhibit HMGB1 [10].
IL-33, in contrast, reflects more the activation of the immune system and cell damage and bridges innate and adaptive immunity. The Th2-skewed cytokine profile in CF has been linked to a deficiency in producing IFN-γ [11]. This has also been observed in response to IL-33 [6]. Given the fact that Th2-type cells upregulate the IL-33 receptor ST2, a good responsiveness to IL-33 may further add to pathophysiology in CF. Furthermore, IL-33 mediates neutrophil influx [5], a characteristic feature in CF. Exposure to Pseudomonas aeruginosa has been shown to induce IL-33 but not HMGB1 mRNA. This is in line with our data, and also with Hartl et al. [12], linking chronic P. aeruginosa infection with augmented Th2 responses. In addition, a linkage between IL-33 mRNA expression and the absence of cystic fibrosis transmembrane conductance regulator (CFTR) has been found [5]. This study significantly adds to current knowledge by confirming IL-33 protein in BALF, in the context of CF.
Taken together, IL-33 may significantly contribute to Th2 responses observed in CF lungs. IL-33 augments Th2-driven inflammation and thereby facilitates chronic inflammation in CF lungs, by an inefficient suppression of inflammatory responses. Thus, IL-33 might influence not only CF lung disease but also other CF-related pathologies. The differential roles of IL-33 and HMGB1 could serve as a model for a better understanding of the complex relationship between innate and adaptive immunity.
Footnotes
Support statement: Supported by funds from the Oesterreichische Nationalbank (Anniversary Fund, project number 13846ONB) and the Medical Scientific Fund of the Mayor of the City of Vienna (number 11013).
Conflict of interest: Disclosures can be found alongside the online version of this article at erj.ersjournals.com
- Received March 10, 2014.
- Accepted April 14, 2014.
- ©ERS 2014
References










